| AVICULTURA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Genética y requisitos cambiantes de aminoácido El desarrollo de linajes de broilers de alto rendimiento ( alto índice de carne en la pechuga principalmente) dicta una necesidad de evaluar los requisitos de nutrientes de broilers desde una perspectiva diferente. Estos linajes de alto rendimiento pueden ser cabalmente alimentados con raciones que pueden tener el equivalente a 25% proteína cruda o más durante el proceso de selección de raza. Esto ha sido práctica común por parte de algunas compañías principales de reproducción por varios años. Sin embargo, no ha habido un ajuste proporcional en los planes de nutrición a nivel industrial. Por lo tanto, algunos broilers comerciales actuales reciben un consumo diario marginal de proteína. Esto puede contribuir a la uniformidad de problemas de crecimiento, rendimiento de carcasa menor a lo esperado, y problemas en las patas. Tenemos evidencia clara que la uniformidad en el crecimiento de los broilers es una función de la proteína en la dieta, especialmente durante los períodos de inicio y crecimiento. Recientemente hemos completado estudios que sugieren que niveles reducidos de aminoácidos esenciales ( específicamente metionina más cistina y lisina) en los períodos de inicio y crecimiento resultan en rendimiento de carne de pechuga reducido a un peso corporal similar. Concentraciones de dieta aceptables de lisina ( expresadas como % de dieta por Kcal ME por gramo de ración) para las raciones de inicio, crecimiento y acabado de linajes de alto rendimiento parecen estar en el rango de 0.397%, 0.386% y 0.245%. De igual modo, la metionina más cistina debería ser aproximadamente 0.310%,0.285% y 0.202% respectivamente para raciones de inicio, de crecimiento y acabado. Para una ración de 3.00 kcal ME /g, el nivel de lisina en la alimentación de inicio será 0.397 x3= 1.191%. Obviamente, es necesario seleccionar el nivel correcto de energía metabolizable en la ración para maximizar el crecimiento. Esto probablemente estará en el rango de 3.2 kcal ME/g ración. Ventilación durante el estrés por calor La alta temperatura siempre ha sido considerada una amenaza para la avicultura pero la combinación de alta temperatura y de alta humedad ha recibido menos atención por parte de la investigación. La influencia de la humedad relativa sobre la pérdida de calor por evaporación fue estudiada por Romijn y Lokhorst (1966) y se muestra a continuación;
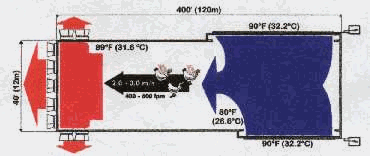
Estos datos son significativos porque muestran que el ave puede aumentar la pérdida de calor (por ejemplo convección) y el almacenamiento de calor cuando la pérdida de calor por evaporación es limitada debido a la alta humedad relativa. Estos datos también sugieren que el emplumado pobre que puede causar ascitis en clima frío puede ser beneficioso en climas calurosos, particularmente en ambientes calurosos, húmedos. El aspecto negativo de un pobre emplume en tales situaciones es un aumento de lesiones de piel (“quemaduras por amoníaco”) debido al contacto de la piel con la cama húmeda. La cama húmeda generalmente se asocia con una mayor excreción urinaria durante el estrés por calor. Las condiciones de la cama pueden mantenerse a niveles aceptables con un mayor flujo de aire en el galpón. Este beneficio aumenta por dos motivos. Primero el mayor flujo de aire elimina el calor del cuerpo eficazmente (ej. pérdida de calor por convección) y reduce la necesidad del ave de aumentar la pérdida de calor por evaporación y urinaria. Segundo, un mayor flujo de aire elimina la humedad cuando el aire fluye sobre la superficie de la cama. El efecto neto en cualquier caso es una temperatura corporal menor que ayuda al consumo de alimento necesario para el crecimiento en un ambiente de galpón mejor (seco). Velocidades de aire más altas pueden obtenerse en ya sea galpones con ventilación por “túnel” (figura 1) o galpones equipados con ventiladores dirigidos hacia abajo ( el opuesto de aquel mostrado en la figura 2). El último puede usarse en casi cualquier estructura de galpón a un costo relativamente bajo. Tales ventiladores se están usando exitosamente en América Latina actualmente. La necesidad de usar ventilación forzada está en función del peso corporal, tasa de crecimiento y plumaje. Generalmente, esto significa que la velocidad del aire necesitará empezar a aumentarse después de los 28 días de edad y posiblemente hasta tan tarde como los 35 días de edad dependiendo de cuándo las aves completan su emplumado y se tornan completamente aisladas. La excesiva velocidad del aire prematuramente aumentará el consumo de alimento sin un aumento asociado en la tasa de crecimiento ( es decir, conversión alimentaria más pobre debido a la excesiva pérdida de calor). Ventilación por túnel
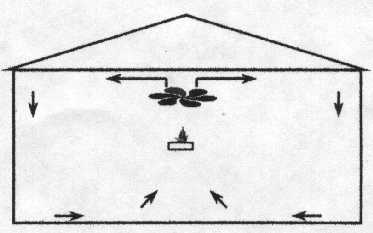
Figura 1. Galpón con ventilación por “túnel” típica. Ascitis La ascitis es aún hoy una preocupación en la producción de broilers. Parece haber una relación estacional con la incidencia de ascitis. La incidencia más baja aparece en los meses más calurosos y la incidencia más alta en los meses más fríos. Es razonable asumir que las diferencias mayores estacionales son temperaturas del ambiente, ventilación y condiciones de galpón asociadas. El amoníaco excesivo contribuye a la ascitis. El rápido secado de la excreta es la clave para controlar la producción de amoníaco, ya que el proceso de secado mata la bacteria que produce amoníaco. Un manejo sugerido es mover el aire caliente del techo del galpón hacia el piso sin una velocidad de aire excesiva a nivel ave. Un arreglo exitoso se ve en la figura 2. Ventiladores a alta velocidad soplando hacia arriba moverán el aire del techo caliente hacia el piso y secarán la excreta cuando ésta se mueve por encima de la cama. La velocidad del aire al nivel del ave es relativamente lenta y no causa amontonamiento de las aves.
Figura 2. Ventiladores dirigidos verticalmente para optimizar las condiciones de crianza.La causa más comúnmente reconocida de ascitis es la rápida tasa de crecimiento. El retardar la tasa de crecimiento (Arce et al., 1990) puede reducir la incidencia de ascitis. Físicamente limitando el acceso al alimento ha mostrado reducir la incidencia de ascitis en proporción al grado de restricción. De igual modo, la alimentación de día alternado (saltear un día) redujo la mortalidad por ascitis (Arce et al., 1990) pero el efecto fue levemente más evidente cuando la restricción se aplicó desde los 22 a los 28 días en lugar de antes de los 22 días (tabla 1). TABLA 1. Desempeño del broiler a los 51 días de edad a una altitud de 1940 metros con influencia de restricción de alimento.
ab Medias que no tienen los mismos exponentes tuvieron diferencia significativa ( P£ .05). Fuente: Arce et al., 1990. Programas de alimentación para el control del síndrome ascíticos. Memorias XV Convención Nacional de la Asociación Nacional de Especialistas en Ciencias Avícolas. Cancún, Q. R., México, pp. 169-177. El hecho de que el clima frío aumenta el consumo de alimento es innegable. El alimento adicional se necesita para la producción de calor, pero tiene el efecto de mantener o aumentar la tasa de crecimiento. Un factor conflictivo es el pobre emplume de los linajes de emplume lento. Mientras que las hembras están generalmente casi totalmente emplumadas a los 21 días de edad, los machos de emplume lento tienen una gran cantidad de área corporal sin cubrir hasta los 35 días de edad. Parece que el frío tiene su mayor impacto cuando las temperaturas de crianza son típicamente bajadas después de los 21 días de edad. Los machos están relativamente sin plumas desde aproximadamente los 21 a 35 días de edad y parecen ser más sensibles al desafío térmico, aunque una exposición más temprana al frío tenga algo de efecto. Esto sugiere que debe prestarse mucho cuidado al mantenimiento de las temperaturas adecuadas para los machos de lento emplume. Una inadecuada proteína en la dieta, aminoácidos sulfurados, arginina y sodio parecen retardar el desarrollo de las plumas. Un espacio de piso insuficiente también contribuye a un pobre emplume y a la eficiencia alimentaria como se ve en la tabla 2. Tabla 2. Efecto de la densidad en la conversión alimentaria (CA) y calidad de la pluma del broiler.
Fuente: Dr. Nick Dorko, Arbor Acres Farm, Inc., Glastonbury, CT, USA. En la estación fría del año o cuando no pueden mantenerse temperaturas ideales, puede ser económicamente factible reducir la tasa de crecimiento de los broilers restringiendo el alimento, ya sea usando restricción física o /y un programa de iluminación. Reduciendo la tasa de crecimiento se reduce por igual la necesidad de oxígeno y se reduce el ritmo del trabajo del corazón. El sodio y el cloro en el alimento no deben ser excesivos si las temperaturas del galpón no son óptimas. Puede usarse bicarbonato de sodio para reemplazar parte del cloruro de sodio para satisfacer los requerimientos mínimos del NRC ( 1994) de sodio y de cloro de 0.20% durante las primeras 3 semanas de edad y 0.15% subsiguientemente. El uso de bicarbonato de sodio también aumenta la relación catión : anión ( balance electrolítico) expresada en miliequivalentes/ Kg de alimento. Un balance electrolítico más positivo ayuda a que los riñones del ave reduzcan el excesivo ácido carbónico que tiende a acumularse cuando la oxigenación de la sangre es pobre. De igual manera, un vasodilatador debe reducir la presión sanguínea y la incidencia de ascitis. El óxido nítrico (ON) es un vasodilatador natural producido a partir de la arginina. Suplementar la dieta con L-arginina redujo la incidencia de ascitis (Wideman et al.,1995b). Por analogía, puede ser posible que las dietas bajas en arginina contribuyan a la incidencia de ascitis. Es bien conocida la interacción de la arginina y la lisina en las dietas. Recientemente se ha encontrado que la absorción de arginina se reduce en condiciones de altas temperaturas (Brake et al.,1994ab) como aquellas normalmente usadas durante la crianza. La respiración pastosa observada a menudo cuando se crían polluelos a altas temperaturas puede ser reducida mediante la suplementación alimentaria con un ingrediente alto en arginina (J. Brake, datos no publicados). Por lo tanto, una adecuada relación arginina:lisina en la ración puede ser necesaria si la crianza se realiza bajo temperaturas cálidas. La tabla 3 muestra las proporciones de arginina:lisina presentes en ingredientes comunes. La formulación exacta de raciones difiere ampliamente pero es de interés notar que las proporciones de arginina:lisina óptimas sugeridas para el máximo crecimiento del broiler son 1.14, 1.10 y 1.18 durante los períodos de 0 a 3 semanas, 3 a 6 semanas y 6 a 8 semanas respectivamente (NRC, 1994). Una reducción en el consumo relativo de arginina parece ocurrir típicamente aproximadamente en la etapa de mayor sensibilidad al frío o mayor tasa de crecimiento en relación a la incidencia de ascitis. Debe tenerse cuidado en la formulación de estas raciones en estos períodos. TABLA 3. Contenido de arginina y lisina en ingredientes comunes.
Fuente: National Research Council, 1994. La tabla de abajo muestra el efecto beneficioso sobre la ascitis de una substitución parcial de pasta de algodón alta en arginina (PA) por harina de soja (HS) (J.D. Garlich, datos no publicados). Basado en estos datos, puede esperarse que una fuente de proteína baja en arginina, tal como harina de pescado, puede contribuir a la ascitis.
PA: 4% de dietas de inicio y 8% de dietas de crianza. Estrategias de nutrición durante el estrés por calor. El manejo del estrés por calor continúa desafiando a la industria del broiler. Se ha sugerido a menudo que las raciones de verano deberían contener menores niveles de proteína cruda complementados con amioácidos esenciales para reducir el incremento de calor de la ración. Sin embargo, el excelente desempeño en climas calurosos y húmedos con raciones altas en proteínas puede a menudo observarse bajo condiciones comerciales (observaciones personales). Es tentador considerar la posibilidad que la ración ideal de aminoácidos podría ser alterada durante el estrés por calor. Alleman y LeClercq (1997) redujeron la proteína cruda de las raciones de crecimiento de broilers de 19.86 a 16.02 % y suplementaron la ración de menor proteína con lisina, treonina, valina y arginina. Estas raciones se alimentaron desde los 23 a 44 días de edad mientras las aves eran expuestas ya sea a 22ºC o a 32ºC. La ración de menor proteína produjo aves con más grasa abdominal, menos músculo del pecho, y conversión alimentaria ligeramente más pobre. El crecimiento fue similar a 22ºC pero la ración de menor proteína redujo el crecimiento a 32ºC. Estos autores concluyeron que la estrategia de disminuir la proteína mientras se agregaban aminoácidos esenciales para combatir el estrés por calor era errada. Ellos sugirieron que el balance de aminoácidos que usaron fue incorrecto para este propósito. La proporción de Arginina:Lisina era 1.24 y 1.07 en las raciones de proteína normal y baja, respectivamente. El objetivo de un programa de investigación entre la Universidad de Carolina del Norte y la Universidad de Sydney es delinear los efectos de la ración Arginina:Lisina en la dieta (Arg:Lis) sobre el desempeño del broiler. Nosotros hemos informado recientemente que la proporción dietética Arginina:Lisina es importante para el desempeño durante el estrés por calor. Dependiendo del balance dietético de electrolitos (BDE), especialmente el sodio y cloro, la proporción óptima puede ir de 1.05 a 1.35. En una ración baja en sodio, como la de Alleman y LeClercq (1997) una proporción más alta debe dar mejores resultados durante el estrés por calor. Estos hallazgos son consistentes con observaciones personales que muestran que aves que reciben raciones nutricionalmente densas se desempeñan bien en países tropicales. Ración Arginina: Lisina y electrolitos en la dieta. En el experimento 1 los broilers fueron alimentados con raciones de Arg:Lis de 1.10 o 1.37 a temperaturas cíclicas de 18-26ºc o 25-35ºC desde los 20 a los 47 días de edad. Los broilers crecieron menos y consumieron menos alimento a temperaturas más altas como se esperaba (tabla 4). No se observó ningún efecto significativo de la dieta sobre la ganancia de peso y el consumo de alimento ya sea en dieta de temperatura o sobre la conversión de alimento a temperaturas más bajas. Sin embargo hubo una mejora significativa en la conversión de alimento a temperaturas más altas en broilers alimentados con una ración de 1.37 Arg:Lis. Los broilers con la ración de Arg:Lis de 1.37 tendieron a comer más alimento a temperaturas más bajas y menos alimento a temperaturas más altas que los broilers alimentados con raciones de 1.10 Arg:Lis (tabla 4). En el experimento 2 (tabla 5) los broilers fueron alimentados con dietas con raciones de 1.05, 1.20, 1.34 o 1.49 de Arg:Lis a 31ºC desde los 28 a los 49 días de edad con niveles de NaCl de 1.2 o 2.4 g/kg de dieta. Los broilers alimentados con las dietas más bajas en NaCl mostraron mejoras consistentes en la conversión de alimento y ganancia corporal aumentando la ración de Arg:Lis, lo que no se vio en los broilers alimentados con dietas conteniendo mayores niveles de NaCl (tabla 5). Se observó una interacción significativa Arg:Lis por NaCl para la conversión del alimento así también como un efecto significativo Arg:Lis para la conversión del alimento. En el experimento 3 (tabla 6) los broilers fueron alimentados con raciones de Arg:Lis de 0.94, 1.08, 1.25 o1.39 desde los 42 a 55 días de edad en ambientes térmico neutrales o de estrés por calor. Los broilers sujetos a temperaturas elevadas nuevamente exhibieron una reducida ganancia corporal y consumo de alimentos como se esperaba (tabla 6). Hubo una interacción significativa ambiente térmico x dieta para la conversión alimentaria que se debió a una mejor conversión alimentaria con una ración mayor de Arg: Lis en el ambiente caluroso que no fue aparente en el ambiente térmiconeutral. Los beneficios de la ganancia de peso, consumo de alimento y conversión de alimento que resultaron de una ración mayor en Arg:Lis a 31ºC no fueron evidentes a un nivel más alto de NaCl en la dieta en el experimento 2 (tabla 5). Hubo una interacción significativa entre NaCl en la dieta y la ración de Arg: Lis para la conversión de alimento debido a una mejorada conversión de alimento en la ración 1.05 en la presencia de mayor NaCl y una mejorada conversión de alimento en la ración 1.49 en la presencia de NaCl más bajo (tabla 5). El análisis estadístico indicó que la ración Arg:Lis óptima fue reducida de 1.34 con 1.2 gr NaCl/kg a la ración más baja examinada (1.05) con 2.4 g NaCl/kg. También hubo interacción significativa entre el ambiente (TN vs HS) y la ración Arg:Lis para la conversión del alimento en el experimento 3 (tabla 6). En el ambiente térmiconeutro se encontró la mejor conversión de alimento en una ración Arg: Lis de 0.94 mientras que el mejor en el ambiente de estrés por calor fue, estadísticamente, no menor que 1.08. Ambas raciones caen bajo la ración óptima de 1.18 calculada por el National Research Council (1994) para los broilers de 6 a 8 semanas. Las dietas que se usaron en este estudio contenían una alta concentración de NaCl típica de las dietas de USA y más altas que aquellas que se usaron en el experimento 1 (tabla 4). Los datos de los experimentos 2 y 3 sugieren que las dietas de broilers que contienen altas concentraciones de NaCl tendrán estrechas raciones óptimas de Arg:Lis pero la ración óptima de Arg:Lis aún varía entre condiciones TN y HS (Térmiconeutral vs estrés por calor). Se han hecho estudios para determinar el efecto de un agudo estrés por calor sobre el consumo de lisina y arginina cuando ambas estaban presentes en condiciones equimolares (1.5 mol/mL). Los datos indicaron que hubo una reducción significativa en el consumo sodio independiente de arginina por el epitelio intestinal de aves estresadas por calor que se reflejó en una reducción significativa del consumo total de arginina. Aunque las mediciones in vitro del consumo de aminoácidos desde la traquea intestinal han limitado el valor cuando se considera en término de los muchos factores que afectan el consumo y utilización de aminoácidos en el ave viva, los resultados ayudan a proveer una explicación posible para las respuestas observadas en los estudios in vivo. Herzberg et.al. (1971) observó que el consumo de lisina y arginina era moderadamente sodio dependiente, mostrando aproximadamente el 50% de reducción en la ración inicial en la ausencia de sodio. De manera muy interesante, el aumento de acumulación en lisina que se observó en el intestino de aves estresadas por calor pareció ser exclusivamente sodio dependiente. La suplementación de sodio de una dieta alta en lisina (es decir reducida ración arg:lis) en combinación con un ión metabolizable tal como el bicarbonato ha mostrado mejorar el antagonismo arg:lis de manera similar a la arginina adicional (O’Dell y Savage, 1966). Los datos de los experimentos 2 y 3 (tablas 5 y 6) sugieren que el NaCl puede tener un efecto similar. El experimento 4 examinó el efecto de 0 o 16 gr/Kg de bicarbonato de sodio en la presencia de raciones de arg:lis de 1.05, 1.15, 1.25 y 1.35 a 21ºC y 31ºC desde los 21 a 41 días de edad (tabla 7). La inspección de los medios de interacción reveló grandes aumentos en el consumo de alimento como resultado de la adición de NaHCO3 a las dietas con raciones de arg:lis de 1.25 y 1.35 a 21ºC y con raciones de arg:lis de 1.15 y 1.35 a 31ºC. Se observaron mejoras numéricas correspondientes en la ganancia de peso corporal a 31ºC pero no a 21ºC. En la presencia de NaHCO3 hubo una mejora dosis relacionada en el consumo del alimento con una ración arg:lis mayor de 1.5 a 1.35 a 21ºC pero una respuesta más amplia maximizada a 1.15 a 1.25 a 31ºC. Aunque no hubo una mejora correspondiente en la ganancia de peso corporal a los 21ºC resultante de la combinación de NaHCO3 y la ración de arg:lis, se observaron mejoras en la ganancia del peso corporal en las raciones arg:lis de 1.15 y 1.25 a 31ºC. Sólo hubo mortandad en aves a los 31ºC. El modelo de mortandad para aves que no recibieron NaHCO3 fue 15,5, 5 y 0%, respectivamente, sobre las dietas con raciones de arg:lis de 1.05, 1.15, 1.25 y 1.35. La mortandad para aves que recibieron NaHCO3 fueron 5% sobre las dietas con raciones de arg:lis de 1.05, 1.15 y 1.35. NaHCO no parece afectar consistentemente el ph en sangre en broilers estresados por calor (Teeter et. al. 1985; Branton et. al. 1986). Las mejoras de desempeño obtenidas con este suplemento aparecen más frecuentemente en la presencia de raciones amplias de arg:lis ( Balnave y Brake, datos no publicados). Un posible modo por el cual NaHCo pudiera actuar a altas temperaturas con dietas con raciones altas en arg:lis sería suministrar iones de bicarbonato para amortiguar las concentraciones mayores de arginina, un catión altamente cargado, en la sangre. El pKa del grupo de guanidina de arginina es 12.48 comparado con un pKa para el grupo amino epsilon de lisina de 10.53. Esto divorciaría el mecanismo de acción de NaHCO3 de la traquea digestiva, y al mismo tiempo, confirmaría previas sugerencias de un requisito nutricional de bicarbonato en altas temperaturas. Los resultados de nuestros experimentos con broilers sugieren una necesidad de re-evaluación de la importancia de una adecuada nutrición de arginina relativa a otros aminoácidos como la lisina. La nutrición de arginina es especialmente importante durante el tiempo caluroso ya que su requisito metabólico aumenta. Sin importar el mecanismo, el riesgo de una falta de equilibrio de arginina es aparente durante el clima caluroso. Una corrección apropiada de esta falta de equilibrio aparente por formulación de ingrediente o suplementación de arginina puede resultar en una mejor conversión alimentaria y una tasa de mortalidad más baja. Es necesaria una mayor investigación con respecto a la nutrición de arginina para las aves para entender mejor su rol durante condiciones de crecimiento estresantes. Ración arginina:lisina y actividad de la metionina. Existe considerable controversia con respecto a las eficacias relativas de DL-metionina (DLM) y análoga hydroxi metionina (MHA) compuestos. Esto ha llevado a la sugerencia que la composición de la dieta en las cuales se incorpora las fuentes de metionina pueden influenciar sus eficacias relativas para productos MHA tales como 2-hydroxy-4 (metylthio) ácido butanoico (HMB). Una revisión que hemos hecho del balance aminoácido en las dietas usadas en estudios de eficacia previos sugiere que la ración arg:lis puede tener una influencia importante sobre la eficacia relativa de DLM y HMB. El experimento 5 (tabla 8) fue diseñado como un estudio 3 x 3 de 3 raciones arg:lis (1.03, 1.20 y 1.34) por 3 fuentes de metionina (dieta básica no suplementada, dieta básica + HMB y dieta básica + DLM). La ración arg:lis de la dieta básica limitante en metionina (0.67 % TSAA) dada desde los 21 a 48 días fue reducida por la inclusión de harina de pescado en la dieta; la ración arg:lis de esta dieta básica fue 1.03. La inspección de la tabla 8 muestra que aumentando la ración arg:lis de 1.03 a 1.34 redujo el consumo de alimento desde los 21 a 48 días de edad de aves dadas HMB en sólo 0.7% mientras que hubieron reducciones de 4.6% en el consumo de alimento en aves dadas el suplemento DLM en estas edades. Brake et. al. (1998) ( experimento 2 tabla 5 en este trabajo) informó que la ración óptima arg:lis para broilers alimentados con dietas conteniendo DLM a altas temperaturas decrecían desde aproximadamente 1.34 a 1.05 o menos cuando el cloro de sodio (NaCl) se usaba para aumentar las concentraciones de sodio y cloro de 0.17% y 0.20%, respectivamente a 0.22 y 0.27%, respectivamente. En concentraciones de cloro y sodio de 0.19 y 0.29 %, respectivamente, la ración óptima de arg:lis aproximó 1.03 sobre las dietas DLM en el experimento 5 (tabla 8). Esto sugiere que el componente de cloro NaCl fue más responsable por bajar la ración óptima de arg:lis en aves alimentadas con DLM en el experimento 2 (tabla 5) que el componente de sodio como se sugirió anteriormente. Estas repuestas son consistentes con altas concentraciones de cloro mejorando la disponibilidad de arginina relativa a lisina, por lo tanto reduciendo la ración arg:lis óptima. También es interesante notar que HMB no reaccionó de igual manera que DLM a los cambios en la ración arg:lis a temperaturas elevadas usadas en estos estudios. En el caso de HMB, el consumo de alimento, la ganancia de peso corporal y la conversión alimentaria fueron mantenidas a niveles similares a través de todas las raciones arg:lis en la dieta. No hubo diferencias significativas en el desempeño de los broilers alimentados con suplementos equimolares de DLM y HMB cuando se midieron las tres raciones arg:lis en la dieta usadas en el estudio. Esto es consistente con los resultados de otros estudios de broilers utilizando dietas prácticas. Se han observado anteriormente mejoras similares en la eficacia aparente de HMB relativa a DLM como resultado de aumentar la ración arg:lis de una dieta práctica (Thomas et. al, 1991). Los resultados actuales sugieren que el efecto de la ración arg:lis en la dieta está resuelto a través de respuestas en el consumo del alimento y sugiere que la elección de una fuente de actividad metionina debería estar acompañada de un conocimiento de la ración arg:lis en la dieta así también como de electrolitos en la dieta. TABLA 4 Respuestas de producción a la arginina en la dieta entre los 20 y 47 días de edad en experimento 1 (Brake et. al. 1998. British Poultry Science 39: 639-647)
ab medios sin sobrescritos comunes son significativamente diferentes en P <0.05. 1 Número de jaulas de duplicado de 6 aves. 2 Análisis fue un lote -dividido sin duplicación para el lote principal de temperatura y sin testeo estadístico de los efectos del lote principal. TABLA 5 Respuestas de producción a la suplementación básica libre en L-arginina entre los 28 y 49 días de edad a 31ºC en experimento 2 (Brake et. al. 1998. British Poultry Science 39: 639- 647)
1 Número de jaulas de duplicación de 5 aves TABLA 6 Respuestas de producción a la suplementación básica libre en L-arginina entre los 42 y 55 días de edad en ambientes térmiconeutrales (TN) o con estrés por calor (HS) en experimento 3 (Brake et. al. 1998. British Poultry Science 39: 639-647) Ganancia Consumo Conversión n1 peso alimentario alimentaria (g alimento/ g ganancia) (g) (g) (g) Efectos Principales2 Ambiente TN 1027 2598 2.531 HS 790 2378 3.007 Arg:lis 0.94 8 883ª 2534ª 2.871ª 1.08 8 907ª 2531ª 2.794ª 1.25 8 912ª 2499ª 2.743ª 1.39 8 932ª 2498ª 2.678ª SEM 78 218 0.083 Interacción Ambiente Arg:lis TN 0.94 4 1065ª 2588ª 2.427e 1.08 4 1021ª 2624ª 2.571d 1.25 4 1003ª 2588ª 2.579d 1.39 4 1017ª 2583ª 2.537de HS 0.94 4 701ª 2320ª 3.308ª 1.08 4 793ª 2387ª 3.011bc 1.25 4 820ª 2370ª 2.893bc 1.39 4 846ª 2386ª 2.818c SEM 112 308 0.119 a-e los medios sin sobrescritos comunes son significativamente diferentes a P<0.05. 1 número de corrales de duplicación de aproximadamente 20 aves. 2 El análisis fue un lote-dividido sin suplicación para el lote principal de temperatura y sin testeo estadístico de los efectos del lote principal.
TABLA 7 Respuestas de producción entre los 21 y 41 días de edad de broilers alimentados con raciones variantes de arginina:lisina (arg:lis) sin y con bicarbonato de sodio (NaHCO3) a 21ºC y 31ºC en experimento 4 (Balnave y Brake, 1999. Australian J. Agric. Research 50: 425-430)
Ganancia Consumo Conversión n1 peso 2 alimentario2 alimentaria 2 (g alimento/ g ganancia) (g) (g) (g) Medios de efecto principalTemperatura (ºC) 21 32 1129ª 2486ª 2.20ª 31 32 1001b 2133b 2.13b SEM 10.0 22.9 0.019 l.s.d.(P=0.05) 28.5 65.2 0.054 Arg:lis 1.05 16 1034 2234b 2.16 1.15 16 1080 2305ab 2.14 1.25 16 1075 2337ª 2.17 1.35 16 1072 2361ª 2.20 SEM 14.2 32.4 0.027 l.s.d. (P=1.05) 92.2 NaHCO3 (gr/Kg) 0 32 1061 2259b 2.13b 16 32 1070 2360ª 2.20ª SEM 10.0 22.9 0.019 l.s.d. (P=0.05) 65.2 0.054 Medios de interacción Temp. Arg:lis NaHCO3 (ºC) (gr/Kg) 21 1.05 0 4 1124 2425 2.16 21 1.15 0 4 1142 2409 2.11 21 1.25 0 4 1159 2458 2.12 21 1.35 0 4 1110 2435 2.20 21 1.05 16 4 1103 2444 2.22 21 1.15 16 4 1133 2482 2.19 21 1.25 16 4 1110 2601 2.34 21 1.35 16 4 1155 2636 2.28 31 1.05 0 4 971 2016 2.08 31 1.15 0 4 980 2075 2.12 31 1.25 0 4 988 2077 2.10 31 1.35 0 4 1011 2179 2.17 31 1.05 16 4 938 2053 2.19 31 1.15 16 4 1064 2255 2.12 31 1.25 16 4 1044 2214 2.12 31 1.35 16 4 1011 2194 2.17 SEM 28.3 64.8 0.053 1 Número de corrales de duplicación de 5 aves cada uno 2 Valores dentro de una medida seguidos por la misma letra no son significativamente diferentes (P>0,05)
TABLA 8. Consumo de alimento, ganancia de peso corporal y conversión alimentaria de broilers de 21-48 días de edad a 32ºC alimentados con suplementos de 0.26% Alimet TM (HMB) y 0.23% DL-metionina (DLM) en dietas conteniendo raciones de Arg:Lis de 1.03, 1.20 y 1.34 en Experimento 5 ( Balnave et al., 1999. J. Applied Poultry Science).
Consumo Ganancia Conversión alimentario peso alimentaria
(g alimento/ g ganancia)
Ración Arg:lis 1.03 3262 1732 1.89 1.20 3260 1747 1.87 1.34 3267 1751 1.87 Probabilidad NS NS NS Suplemento Ninguno 3221 1643b 1.97ª HMB 3317 1816ª 1.83b DLM 3251 1772ª 1.84b Probabilidad NS 0.0006(31.3)1 0.0001(0.014)
Interacción Suplemento Arg:Lis Ninguno 1.03 3116 1559b 2.00a Ninguno 1.20 3261 1681ab 1.94ª Ninguno 1.34 3286 1690ab 1.96ª HMB 1.03 3333 1813ª 1.84b HMB 1.20 3306 1804ª 1.84b HMB 1.34 3311 1831ª 1.81b DLM 1.03 33382 1824ª3 1.83b DLM 1.20 3233 1769ª 1.83b DLM 1.34 3183 1721ª 1.83b Probabilidad NS 0.003(47.7) 0.0001(0.025) ab medios sin sobrescritos similares no difieren significativamente ( P<0.05). 1 SEM dado entre paréntesis donde se obtuvo significancia. 2 Hubo una correlación negativa (P=0.083, R2=0.983) para consumo de alimento con una ración mayor de A:L en la presencia de DLM solamente. 3. Hubo una correlación negativa significativa (P= 0.004, R2=0.999) para una ganancia de peso con una ración mayor de A:L en la presencia de DLM solamente.
Bibliografía ALLEMAN, F. & LECLERCQ, B. (1997) Effect of dietary protein and environmental temperature on growth performance and water consumption of male broiler chickens. British Poultry Science 38:607-610. BALNAVE, D. & BRAKE, J. (1999) Responses of broilers to sodium bicarbonate supplementation of diets containing varying arginine : lysine ratios. Aust.J.Agric.Res.50:425-430. BALNAVE, D. , HAYAT,J. & BRAKE, J. (1999) Dietary arginine:lysine ratio and methionine activity at elevated environmental temperatures. J. Appl. Poultry Reserach 8:1-9. BRAKE, J. , BALNAVE, D. & DIBNER, J.J. (1994 a) Wide Arg:Lys ratio ameliorates effect of heat stress in broilers. Poultry Science, 73 (Suppl. 1) : 74 (abs). BRAKE, J., FERKET, P., GRIMES, J., BALNAVE, D., GORMAN, I. & DIBNER,J.J. (1994 b). Optimum arginine lysine ratio changes in hot weather. Proceedings 21st Annual Carolina Poultry Nutrition Conference. Charlotte, N.C. pp. 82-104 BRAKE,J., BALNAVE, D. & DIBNER, J.J. (1998). Optimum dietary arginine:lysine ratio for broiler chickens is altered during heat stress in association with changes in intestinal uptake and dietary sodium chloride. British Poultry Science 39:639-647. BRANTON, S.L., REECE, F.N. & DEATON, J.W. (1986) Use of ammonium chloride and sodium bicarbonate in acute heat exposure of broilers. Poultry Science 65, 1659-1663. HERZBERG, G.R., SHREERING, S. & LERNER,J. (1971) Cationic amino acid transport in chicken small intestine. Comparative Biochemistry and Physiology, 40 A:229-247. NATIONAL RESEARCH COUNCIL (1994). Nutrient requirements of poultry. O’DELL, B. L. & SAVAGE, J.E. (1966) Arginine:lysine antagonism in the chick and its relationship to dietary cations. Journal of Nutrition, 90:364-370. ROMIJN, C. & LOKHORST, M.O. (1996) Pages 211-227 in: Physiology of the domestic fowl, (Horton-Smith, C. and Amoroso, E. C., Eds.), Oliver and Boyd, Edinburgh. TEETER, R. G., SMITH, M.O.,OWENS, F. N., ARP, S.C., SANGIAH, S. & BREAZILE, J. E. (1985). Chronic heat stress and respiratory alkalosis: occurrence treatment in broiler chicks. Poultry Science 64, 1060-4. THOMAS, O. P., C. TAMPLIN, S. D. CRISSEY, E. BOSSARD, & A. ZUCKERMAN, 1991. An evaluation of methionine hydroxy analog free acid using a non-linear (exponential) bioassay. Poultry Science 70: 605-610. WIDEMAN, R. F., Jr., Y. KOCHERA KIRBY,M. ISMAIL, W. B. BOTTJE, R. W. MOORE, & R.C. VARDEMAN, 1995b. Supplemental L-Arginine attenuates pulmonary hypertension syndrome (ascites) in broilers. Poultry Science 74: 323-330.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||